Ричард
Докинз
Самое
грандиозное
шоу на Земле:
доказательства
эволюции

Глава
8
Ты
сделал это
сам – за
девять
месяцев
Однажды
после
публичной
лекции
гениальному
Джону Бердону
Сандерсону
Холдейну
(помимо
прочего –
одному из
трех
архитекторов
современного
неодарвинизма)
бросила вызов
некая дама. К
сожалению,
эта история
передается
из уст в уста,
а Джона Мейнарда
Смита,
который мог
бы вспомнить
точные слова,
с нами больше
нет. Диалог
был примерно
таким.
Дама: Даже
если в
распоряжении
эволюции
были миллиарды
лет, как вы
утверждаете,
я просто не
могу
поверить в
то, что из
одной клетки
может
развиться
сложный
человеческий
организм,
состоящий из
триллионов
клеток,
сложившихся
в кости,
мышцы и нервы,
в сердце,
которое
безостановочно
работает
десятилетиями,
во многие
мили кровеносных
сосудов и
почечных
канальцев, в
мозг,
способный
мыслить и
чувствовать.
Профессор
Холдейн: Но,
мадам, вы
же сами это
проделали. И
у вас ушло на
это всего
девять
месяцев!
Думаю,
что дама была
ошарашена.
Думаю, что сравнение
с парусами,
внезапно
оставленными
ветром, будет
вполне
уместно.
Однако минимум
в одном
аспекте
ответ Холдейна
мог оставить
ее
неудовлетворенной.
Не знаю, задавала
ли она
дополнительные
вопросы, но если
да, то могла
сказать
примерно
следующее:
"Однако
развивающийся
эмбрион
следует заложенным
генетическим
инструкциям.
Вы, профессор,
утверждаете,
что эти
инструкции для
построения
сложного
тела
появились в
результате
эволюции под
действием
естественного
отбора. И в
это мне по
прежнему
сложно
поверить, даже
если
эволюция шла
бы миллиарды
лет".
Вполне
законная
постановка
вопроса. Даже
если было бы
доказано, что
жизнь на Земле
своей
сложностью и
разнообразием
обязана
высшему
разуму, он
все же,
безусловно,
не лепит
живые тела,
как
скульптор из
глины, и не
изготавливает
их, как
плотник, портной
или
производитель
автомобилей,
которые
производят
свою
продукцию.
Да, мы “замечательно
развиты”, но
ни в коем
случае не
“замечательно
сотворены”.
Если Бог и
вмешивается, то
направляя
эмбриональное
развитие, например,
подбором
последовательности
генов. Крылья
никто не
творит:
внутри яйца
они постепенно
развиваются
из зачатков
конечностей.
Бог ни
разу за свою
предположительно
вечную жизнь
не сотворил
ни единого
крылышка. Если
он что-то и
сотворил (с
моей точки
зрения – нет,
но мы сейчас
говорим не об
этом), то это был
эмбриологический
рецепт,
что-то вроде компьютерной
программы,
управляющей
эмбриональным
развитием
крылышка (и
массы других
вещей). Бог,
конечно, мог
бы сказать,
что
придумать
рецепт крыла
не менее сложно,
чем
сотворить
крыло. Но
сейчас я
хочу
заострить
ваше
внимание на отличии
сотворения
крыла или
чего-то подобного
от того, что
происходит
во время
эмбрионального
развития.
Танец
без
хореографа
Ранний
этап
становления
эмбриологии
был отмечен
баталиями
между
сторонниками
преформизма
и эпигенеза.
[Преформизм
(от лат. «заранее
образую») -
учение о
наличии в
половых
клетках материальных
структур,
предопределяющих
развитие
зародыша и
признаки
развивающегося
из него
организма;
Эпигенез (от
греч. «после» и
«рождение») -
учение о
зародышевом
развитии
организмов, в
ходе
которого
происходят
последовательные
новообразования
органов.]
Различия
между этими
доктринами
не всем хорошо
известны,
поэтому я
потрачу
некоторое
время на
разъяснения.
Сторонники
преформизма
были
убеждены в
том, что
яйцеклетка или
сперматозоид
(преформисты
делились на два
лагеря –
“овистов” и “спермистов”)
содержит крошечного
человечка
гомункула.
Все части тела
человечка
уже
находятся на
своих местах,
правильно
сориентированы
друг относительно
друга, и для
развития
необходимо только
увеличение в
размерах
(напоминает надувание
воздушного
шара,
разделенного
на отсеки). Эта
концепция
неизбежно
порождает
ряд вопросов.
Во-первых,
если
следовать ей,
мы приходим к
утверждению,
что ребенок
наследует
все признаки
только
одного из
родителей – отца
(если верить спермистам)
или матери (у
овистов),
хотя все
понимают, что
это не так.
Во-вторых,
преформисты
сталкиваются
с проблемой
упаковки
уменьшающихся
матрешек (гомункулов
внутри
гомункулов) –
если не бесконечно,
то вплоть до
Адама (спермисты)
или Евы
(овисты).
Единственный
способ избежать
этой
регрессии –
предположить
формирование
нового
гомункула в
каждом
поколении
при помощи
полного
“сканирования”
взрослого организма.
Однако и
здесь
проблема:
приобретенные
свойства не
наследуются.
В противном
случае
мальчики,
рожденные в
ортодоксальных
иудейских
семьях, с
рождения
были бы лишены
крайней
плоти, а
посетители
фитнес
клубов, в отличие
от лежебок,
производили
бы на свет младенцев
с “кубиками”
на прессе и с
перекачанной
грудью и
ягодицами.
Сказанного
достаточно,
чтобы понять,
что у преформизма,
по крайней мере в его
изначальной,
“матрешечной”
версии, не
было шансов.
Но возможен
ли вариант
преформизма,
жизнеспособный
в эпоху ДНК? В
принципе да,
хотя и сомнительный.
Учебники
биологии
снова и снова
повторяют:
ДНК – это
“чертеж” для
строительства
тела. Это не
так.
Настоящий
чертеж,
например автомобиля
или здания,
точно
отражает
результат
(учитывая
масштабирование,
естественно).
Следовательно,
любой чертеж
можно воссоздать,
исследовав
конечный
продукт. Создать
чертеж по
зданию
ничуть не
сложнее, чем
возвести
здание по
чертежу –
именно потому,
что они точно
соответствуют
друг другу.
Это даже
проще,
поскольку,
действуя в
обратном направлении,
вам надо
строить, в
то время как
при создании
чертежа –
всего-навсего
провести
замеры и
нарисовать.
Но сколько
ни измеряй
тело
животного,
это не
поможет вам реконструировать
его ДНК.
Поэтому
называть ДНК
чертежом
неверно.
В
принципе,
можно
представить
себе ситуацию,
в которой ДНК
является
чертежом
организма.
Может быть,
на далекой
планете
именно так
устроена
жизнь:
молекула ДНК
представляет
собой
трехмерную
карту,
закодированную
в линейной
последовательности
“буквами”
генетического
кода. Такая
карта была бы
вполне
обратимой.
Идея получения
подробного
молекулярного
“чертежа”
путем
сканирования
организма не
является
абсолютно
невероятной.
И, если бы ДНК
действительно
работала так,
то можно было
бы создать концепцию
неопреформизма,
не вспоминая
о “проблеме
матрешек”.
Правда, мне
не совсем
понятно, как
решалась бы в
таком случае
проблема
наследования
только от
одного
родителя. ДНК
обладает
поразительным
по точности механизмом
объединения
в потомстве
половины
материнской
с половиной
отцовской наследственной
информации.
Но как бы ДНК
справилась с
задачей
объединения
половины карты
отцовского
организма с
половиной карты
материнского?
Впрочем,
неважно: в
любом случае
это
нереально.
Сегодня мы
уже знаем,
что ДНК – это
не чертеж.
Историческая
альтернатива
преформизму –
эпигенез.
Если
преформизм
исходит из
существования
“чертежей”, то
эпигенез
подразумевает
что-то вроде
"рецепта"
или "компьютерной
программы".
Определение,
данное в
Кратком
Оксфордском
словаре английского
языка, звучит
весьма
современно, и
я сомневаюсь,
что
Аристотель,
автор термина
“эпигенез”,
узнал бы его:
"Эпигенез.
Представление
о
зародышевом
развитии
организмов
как процессе,
осуществляющемся
путем последовательной
дифференциации
изначально недифференцированного
целого"
В книге
Льюиса Уолперта
и его коллег
“Принципы
развития”
эпигенез описывается
как идея
поступательного
развития
новых
структур. В
этом смысле
реальность
эпигенеза
очевидна, но,
как обычно,
дьявол
прячется в
деталях. Как
именно идет
прогрессивное
развитие
организма?
Откуда
“изначально
недифференцированному
целому”
известно, как
развиться в
сложный организм,
как не из
чертежа? Одно
очень важное отличие,
которое я бы
хотел
подчеркнуть,
представляет
собой, по
сути,
основное
отличие эпигенеза
от преформизма.
Это разница
между
строительством
по плану и самосборкой.
Мы все знаем,
как строят
дома, как
изготавливают
вещи,
окружающие
нас, и
поэтому
отлично
понимаем, что
такое
строительство
происходит
согласно
плану. Самосборка
менее очевидна
и
распространена,
поэтому ей
придется уделить
больше
внимания.
Можно
сказать, что
в эмбриогенезе
самосборке
принадлежит
роль, которую
в
эволюционном
процессе
играет
естественный
отбор (хотя это,
безусловно,
разные
явления). Оба
за счет
автоматических,
непреднамеренных,
никем не
запланированных
процессов
приводят к
результатам,
которые на
первый
взгляд
кажутся
тщательнейшим
образом
спланированными.
Холдейн,
отвечая на
вопрос,
конечно,
сказал
чистую правду.
Однако он не стал бы
отрицать, что
в развитии
человеческого
тела всего из
одной клетки
есть тайна,
близкая к
чуду, хоть и
не
переходящая
его грань. То,
что развитие
происходит
по
инструкциям,
содержащимся
в ДНК,
приоткрывает
завесу таинственности,
но не
полностью.
Причина
таинственности
проста: нам
сложно
представить,
даже в
теории,
возможность
создания инструкций
[то есть
"рецепта"]
для строительства
тела
способом,
которым оно
как раз и строится
на самом
деле: самосборкой.
На принципе самосборки
основаны
некоторые
компьютерные
программы и
типы
поведения животных.
Предположим,
что мы хотим
смоделировать
стайное
поведение
скворцов. В
стайном
поведении
скворцов
поразительнее
всего то, что
у них нет
хореографа и,
насколько
нам известно,
вожака также.
Каждая птица всего
лишь следует
локальным
правилам.
Число
птиц в
подобных
стаях может
измеряться
тысячами,
однако
столкновений
практически
не бывает.
Еще бы, ведь
на таких
скоростях
любое
столкновение
почти
неизбежно приведет
к ранениям.
Часто можно
наблюдать,
что стая ведет
себя как
единое целое,
согласованно
поворачиваясь
или кружась.
Дополнительное
эстетическое
удовольствие
дает четкий
абрис стаи:
ее плотность
не снижается
от центра к
периферии. Ну
не
удивительно
ли?
Не
стоило бы пытайться
выступать в
роли
хореографа
этого
"балета". Для
подобной
задачи этот
подход
крайне
неудачен. Я
расскажу о
верном способе
составления
подходящего
к этому случаю
алгоритма,
потому что
сами птицы
(их мозги)
почти
наверняка
запрограммированы
так же. А
главное, это
прекрасная
аналогия
эмбрионального
развития.
Вот как
надо
программировать
стайное поведение
скворцов.
Нужно
сосредоточиться
на
программировании
поведения
отдельной птицы.
Встроим в
компьютерного
скворца правила
поведения в
полете и
реагирования
на
присутствие
вблизи
других
скворцов.
Определим, в
какой
степени
птица при
изменении
направления
полета будет
отдавать приоритет
поведению
окружающих особей и в какой –
собственной
инициативе.
Все эти
правила можно
установить
экспериментальным
путем,
наблюдая за
птицами в
природе.
Дадим киберптице
возможность
менять
правила
поведения случайным
образом, но в
определенных
рамках. Теперь,
когда мы
создали
сложную
программу, в
деталях
описывающую
поведение
скворца,
наступает
самый важный
момент. Не
следует
пытаться
программировать
поведение стаи,
как,
возможно,
поступил бы
программист
прошлого.
Вместо этого
мы клонируем
компьютерного
скворца,
создадим
тысячу его
копий, идентичных
или с
небольшими
случайными
вариациями
правил
поведения. И
“выпустим” их
в
компьютерную
среду
обитания,
предоставив
самим себе,
позволив
взаимодействовать
на основании
общих правил.
Если
правила
поведения
одиночной
птицы заданы
удачно, то
тысячи
компьютерных
скворцов,
каждый в виде
точки на
экране,
поведут себя
как реальная
стая. Главное
же здесь вот
что: у нас нет
ни хореографа,
ни дирижера,
ни вожака.
Порядок, организация,
структура
возникают
как побочный
продукт
системы
правил,
исполняемых
локально, а
не глобально.
Именно так
устроено эмбриологическое
развитие: оно
целиком построено
на
соблюдении
локальных
правил на
разных
уровнях, но
главным
образом на
уровне клетки.
Никакого
хореографа.
Никакого
дирижера.
Никакого
централизованного
планирования.
Никакого
архитектора.
Тела
человека,
орла, крота,
дельфина,
гепарда,
леопардовой
лягушки и
ласточки –
все они столь
восхитительно
и точно
собраны, что
кажется
невозможным
то, что гены,
их
программирующие,
не являются
ни чертежом,
ни генпланом.
Тем не менее
это так. Как и
в случае
компьютерных
скворцов, все
создано
отдельными
клетками,
следующими
локальным
правилам.
Прекрасно
спроектированное
тело
возникает
благодаря исполнению
локальных
правил
отдельными клетками,
без какого бы
то ни было
всеобщего плана.
Клетки
эмбриона “танцуют”
друг с другом
как скворцы в
гигантской
стае. Есть,
однако,
важные
различия. В отличие
от скворцов,
клетки
склеены
вместе,
образуют
слои и блоки: их
“стаи”
называются
тканями.
“Танцуя” и
крутясь, они
образуют
трехмерные
формы, вызывая
инвагинацию (впячивание)
тканей в
ответ на
движение
клеток,
разрастание
или сжатие
тканей в
ответ на
изменение
местных схем
клеточного
роста и
смерти. Мне
нравится аналогия,
предложенная
известным
эмбриологом
Льюисом Уолпертом
в книге
“Триумф
зародыша”:
оригами –
японское
искусство
складывания
фигурок из
бумаги.
Кое-что
о развитии
Удивительно
трудно
подыскать
приемлемую аналогию
процесса
развития
живой ткани,
однако можно
найти
частичное
сходство с
отдельными
сторонами
этого
процесса.
Например,
понятие
“рецепт”
гораздо
лучше
отражает суть,
чем “чертеж”, и
я иногда
пользуюсь
первым для
того, чтобы
объяснить,
почему
последнее
неверно. В
отличие от
чертежа,
рецепт необратим.
Следуя
тексту
рецепта
пирога, в
итоге вы
получите
пирог. Но по
готовому
пирогу вы не
восстановите
рецепт, в
то время как,
видя здание,
вполне можно
воспроизвести
чертеж. Дело
в том, что
части здания
и элементы
чертежа
соответствуют
друг другу.
Не считая
очевидных
исключений
вроде вишенки
в центре,
однозначного
соответствия
между
кусочками
пирога и
строчками
рецепта нет.
На
иллюстрации
показаны три
типа вирусов.
Слева – вирус
табачной
мозаики (ВТМ),
который поражает
листья
табака и
других
представителей
семейства
пасленовых
(например, помидоров).
В центре –
аденовирус,
инфицирующий
органы
дыхания
многих
животных, не
исключая нас.
Справа –
бактериофаг
Т4,
который
паразитирует
на бактериях.
Он выглядит
как лунный
модуль, да и
ведет себя
примерно так
же: приземляется
на
поверхность
бактерии (она
гораздо
крупнее его),
приседает на
своих паучьих
ножках и
запускает
вглубь
клетки зонд,
пробуравливая
ее стенку.
После этих
операций
бактериофагу
остается
внедрить в клетку
свою ДНК.
Вирусная ДНК
захватывает
бактериальный
белок
синтезирующий
аппарат, заставляя
его работать
на себя – воспроизводить
новые вирусы.
Два других вируса,
изображенные
на
иллюстрации,
ведут себя
сходным
образом.
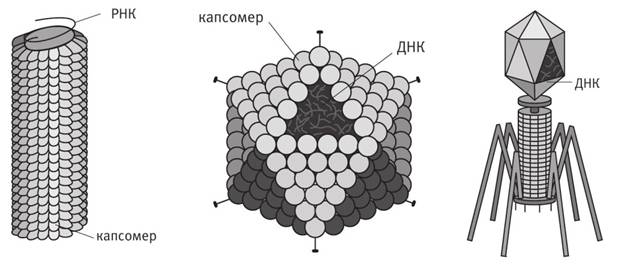
То, что вы
здесь видите,
большей
частью представляет
собой
контейнер с
генетическим
материалом, а
у “лунного модуля”
Т4
заметно еще
приспособление
для заражения
клетки
хозяина.
Любопытно,
как строится
этот
белковый
контейнер: в
процессе самосборки.
Каждый вирус
собирается
из
нескольких
предварительно
синтезированных
белковых
молекул.
Каждая
белковая
молекула сворачивается
в третичную
(объемную)
структуру:
здесь
действуют
законы химии,
распоряжающиеся
конкретными
последовательностями
аминокислот.
Свернутые
молекулы
присоединяются
друг к другу,
формируя так
называемую
четвертичную
структуру.
Здесь
вступают в
силу локальные
правила.
Белковые
субъединицы –
капсомеры
–
присоединяются
друг к другу,
как кирпичики
“Лего”.
Заметьте,
насколько
геометрически
совершенна
эта микроконструкция.
Аденовирус
сложен из 252 капсомеров,
изображенных
здесь в виде
шариков. Они
организованы
в икосаэдр.
Икосаэдр –
одно из
совершенных
“Платоновых
тел”, которое
имеет
двадцать треугольных
граней. Капсомеры
собираются в
икосаэдр не
во исполнение
какого либо
плана, а
потому, что
подчиняются
общим
химическим
законам. Так
формируются
кристаллы, и
аденовирус и
в самом деле
можно
назвать крошечным
полым
кристаллом.
“Кристаллизация”
вирусов
является
превосходным
примером самоорганизации,
которую я
считаю
основным
механизмом,
собирающим
воедино
живых существ.
У
бактериофага
Т4
вместилище
для
генетического
материала также
имеет форму
икосаэдра, но
с более сложной
четвертичной
структурой,
включающей
добавочные
белковые
единицы,
также организованные
согласно
правилам,
диктуемым микроусловиями.
Эти
субъединицы
включены во
впрыскивающий
аппарат
бактериофага
и “ноги”,
прикрепленные
к икосаэдру.
В
настоящем
эмбрионе
клетки,
настоящие живые
клетки, все
время
делятся, и
зародышевые
ткани растут.
И растут они
с разной
скоростью на
разных
участках.
Разница
достигается
различиями в
скорости
деления
клеток,
которая регулируется
микроусловиями.
Итак, мы
вернулись к
основополагающему
принципу:
восходящий
алгоритм
превалирует над
нисходящим.
Имеется
целая серия
вариантов
(существенно
более
сложных)
этого
простого
принципа, который
реализуется
на ранних
стадиях
эмбрионального
развития.
Оплодотворенная
яйцеклетка
делится на две.
Затем из двух
клеток
получается
четыре. Деление
продолжается,
но на этой
стадии еще
нет роста,
нет
разбухания.
Все
заканчивается
сферическим
шариком,
состоящим из
множества
клеток, но
размер его
точно такой,
как и у
яйцеклетки.
Этот шарик –
бластула –
полый.
Следующая
стадия –
гаструляция.
Льюис Уолперт
остроумно
высказался о
ней так:
“Самый важный
момент в
вашей жизни –
не рождение,
не свадьба
или смерть, а гаструляция”.
Гаструляция
охватывает
всю поверхность
бластулы и
переиначивает
все вокруг.
Ткани
эмбриона
радикально
перестраиваются.
Обычная
гаструляция
предполагает
впячивание
полого
шарика, то
есть
бластулы,
внутрь таким
образом, что
он
становится
двухслойным
с одним
отверстием
наружу.
Внешняя стенка
гаструлы
называется
эктодермой,
внутренняя
стенка –
энтодермой,
между этими слоями
располагаются
клетки,
слагающиеся в
мезодерму.
Эти три
зачаточных
слоя в итоге
формируют
все части
тела. Так, из
эктодермы получаются
кожа и
нервная
система,
пищеварительную
систему и
другие
внутренние
органы
формирует
энтодерма, а
из мезодермы
образуются
кости и
мышцы.
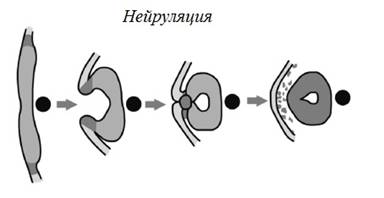
Следующая
стадия
эмбрионального
развития
называется
нейруляцией.
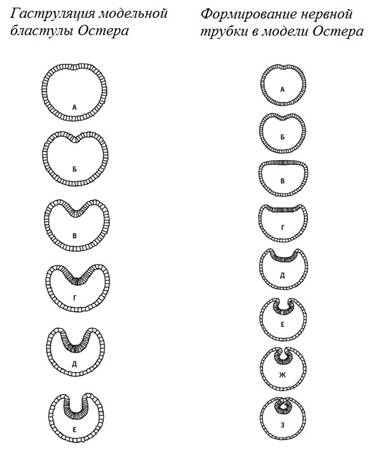
На рисунке
показаны
срезы
эмбриона
амфибии
(лягушки или саламандры),
идущие
сверху вниз и
через центр.
Черный
кружок – это нотохорд,
твердый
стержень,
действующий
как предшественник
позвоночника.
Нотохорд
служит
диагностическим
признаком
хордовых
животных, к
которым
принадлежим
мы и другие
позвоночные,
хотя у нас,
как и у
других позвоночных,
нотохорд
имеется
только в
эмбриональном
состоянии. Во
время
нейруляции,
как и во
время гаструляции,
самым
заметным
событием
является впячивание,
или
инвагинация.
Напомню,
нервная
система образуется
из эктодермы.
Происходит
это так. По
всей спинной
стороне
часть
эктодермы
постепенно впячивается
в
направлении
от переднего
конца к
заднему, при
этом
сворачиваясь
в трубку и
образуя
соединение,
наподобие
застежки
“молнии”;
соединенные
концы
оказываются
между
внешним
слоем и нотохордом.
Эта трубка в
итоге
превращается
в спинной
мозг –
основной
нервный
ствол тела.
Передний его
конец раздувается
и
превращается
в головной
мозг. Из этой
зачаточной
трубки путем
последовательного
клеточного
деления
происходят
все
остальные
нервы.
Не хочу
углубляться
в детали
гаструляции или
нейруляции. Замечу
только, что
они
удивительны
Выше
показано, что
происходит с
клеточным слоем.
Это только
одно из
многих
событий эмбрионального
развития.
Легко
увидеть, насколько
действенной
операцией
может быть инвагинация
(выпячивание).
Она играет
исключительную
роль, как во
время
гаструляции,
так и во
время нейруляции.
Гаструляция
и нейруляция
заканчиваются
на ранних
стадиях
развития
эмбриона и
определяют
его форму.
Инвагинация
и другие трансформации
растущего
организма
происходят и
на этих
ранних
стадиях, и
после, когда
формируются
сложные
органы,
например,
глаза и
сердце. Но,
все же,
принимая во
внимание, что
все эти
операции
выполняются
без внешнего
воздействия,
что приводит
весь этот
механизм в
движение?
Отчасти, как
я отмечал,
простое
разрастание.
Клетки
делятся во
всех
тканевых
листках.
Следовательно,
их площадь
увеличивается,
а так как
деться растущей
ткани некуда,
то она
собирается в складки
или
впячивается.
Но процесс на
самом деле
находится
под жестким
контролем. Его
изучением
занималась
группа
ученых, связанная
с блестящим
биологом
теоретиком
Джорджем Остером
из
Калифорнийского
университета
в Беркли.
Клетки
и скворцы
При
клеточном
моделировании
Остер и его коллеги
следовали
той же
стратегии,
которую мы
упоминали,
говоря о
скворцах.
Вместо того
чтобы задать
программу
поведения
бластуле, они
задали
программу
одной ее
клетке. Затем
ученые в
компьютере
“вырастили”
много клонов
клеток и
проверили,
что
получится,
когда эти клетки
собраны
вместе.
Говорить
“задали поведение
одной клетке”
не совсем
точно, лучше
так: “создали
математическую
модель одной
клетки,
построенную
на основе
известных фактов о
клетке”, но,
все же,
удобнее
пользоваться
первоначальной
простой
формулировкой.
Особенно важно,
что
внутреннее
пространство
клетки связано
пересекающимися
эластичными
нитями микрофиламентами,
способными
сокращаться
наподобие
мускульных
волокон.
Модель Остера
упрощает
клетку до
двухмерного
изображения,
удобного для
представления
на экране компьютера,
оставляя
всего
полдюжины филаментов,
соединяющих
узловые
точки
геометрической
формы
модельной
клетки. В
модели всем микрофиламентам
придаются
определенные
количественные
характеристики,
смысл
которых
хорошо понятен
физикам:
например,
коэффициент
вязкого
затухания
или
константа
упругости. В
действительности
неважно, что
эти характеристики
обозначают;
физики
обожают
использовать
их, когда
имеют дело с
пружинами
или другими
упругими
телами. В
реальной
клетке сокращаться
способны все филаменты,
но для
упрощения
модели Остер
в своей модели
разрешил
сокращаться
только одной
нити из
шести. Если
реалистичный
результат
получится
даже при
отбрасывании
ряда
известных
свойств
клетки, то
уж при
сохранении
большей
части свойств
результат
поведения
усложненной
модельной
системы
должен
только
улучшиться.
Вместо того
чтобы дать микрофиламенту
возможность
сокращаться
как ему
угодно, ученые
встроили
(запрограммировали)
свойство,
которое
присуще
любому
мышечному
волокну: в
ответ на
растяжение
выше
определенного
предельного
значения
нить
сокращается
до меньшей
длины, чем в
равновесном
(не растянутом
и не
сокращенном)
состоянии.
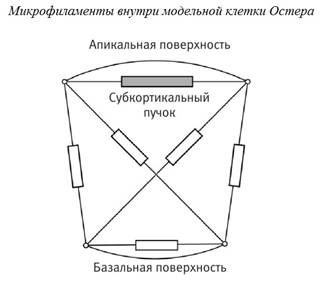
Таким
образом, у
нас есть
сильно
упрощенная
модель
клетки:
двухмерный
абрис, внутри
которого
протянуты
шесть
эластичных
нитей, причем
одна из нитей
способна
активно сокращаться
в ответ на
внешний
сигнал растяжения.
Это первая
стадия
процесса
моделирования.
На второй
стадии Остер
и его коллеги
смоделировали
клон из
нескольких
десятков таких
клеток и
организовали
их в круг, как
будто это
двухмерная
бластула (или
тонкий срез бластулы).
Затем они
взяли и
“потянули”
одну клетку
(запрограммировали
мгновенное
увеличение
натяжения),
чтобы
спровоцировать
сокращение
активного микрофиламента.
То, что
произошло
дальше,
восхитительно:
модельная
бластула
приступила к
гаструляции!
Посмотрите
на шесть
последовательных
кадров,
запечатлевших
это событие.
В обе стороны
от активированной
клетки
распространилась
волна
сокращений, и
клеточный
шарик начал
спонтанно
сгибаться и
впячиваться.
Дальше –
больше. Остер
понизил
пороговое
значение
сокращения активного
микрофиламента.
В результате
начавшееся впячивание
продолжилось,
и в конце
концов выщепилась
“нервная
трубка”.
Важно понять,
что данная модель
– это не
точная
имитация
нейруляции. Но
ведь моделям
и не
полагается
точно повторять
реальные
объекты. А
наша модель
должна
показать,
насколько
просто
имитировать важнейшие
аспекты клеточного
поведения в
раннем
эмбрионе. То,
что даже
простой
двухмерный
кружок из
одинаковых
клеток
спонтанно
отвечает
неким реалистичным
образом на
внешний
стимул, служит
важным
доказательством
того, что
механизм
различных
эмбриональных
процессов на
ранних стадиях
вовсе не
требует
каких-то
сложных решений.
И, конечно же,
нужно
помнить, что
мы имеем дело
с простой
моделью, а не
с простым явлением.
Я
рассказал о
моделях Остера,
чтобы
продемонстрировать
принцип,
лежащий в
основе
взаимодействия
клеток друг с
другом, –
взаимодействия,
в результате
которого они
организуются
в оформленное
тело. Этот
базовый
принцип
справляется
с работой без
всяких
чертежей,
представляющих
идею готовой
конструкции.
Простейшие
операции
построения
раннего
эмбриона уже
были названы:
складывание,
выпячивание
и выщепление,
как в
модельной
бластуле Остера.
Более тонкие
операции
идут на
дальнейших стадиях
эмбрионального
развития.
Так, например,
в ходе
остроумнейших
экспериментов
было
доказано, что
нервные
отростки,
выходящие из
спинного и
головного
мозга, доходят
до места назначения,
до своего
органа, не
следуя заранее
установленному
плану, а за
счет химического
привлечения;
они подобны
псу, находящему
по запаху
суку. Один из
классических
ранних
экспериментов,
превосходно
иллюстрирующих
этот
механизм,
провел
Роджер Сперри
- эмбриолог и
нобелевский
лауреат. Сперри
с коллегами
взяли
головастика
и вырезали у
него со
спинки и с
брюшка
одинаковые
по размеру
квадратики
кожи. А потом
пришили их обратно,
но поменяв
местами:
кусочек со
спинки
оказался на
брюшке, а с
брюшка – на
спинке. Когда
головастик
стал
лягушкой, то
результат
оказался
весьма
забавным, как
это зачастую
бывает в
эмбриологических
экспериментах.
У лягушки на
темной
крапчатой
спинке
красовался
квадратик
кожи с
брюшка, светлый,
как и должно
быть на
брюшке, а на
брюшке выделялся
темный
квадратик
кожи со
спинки. Обычно,
если
пощекотать
лягушку
кисточкой по
спине, она
потрет
раздраженное
место лапой,
будто
сгоняет муху.
Но когда Сперри
пощекотал
светлый
квадратик
кожи на спинке
своей
лягушки, она
потерла
брюшко! А
когда Сперри
щекотал
темный
квадратик на
брюшке, то
лягушка
сгоняла
воображаемую
муху со
спинки.
Согласно
Сперри,
при
нормальном
развитии
эмбриона
происходит
следующее:
аксон –
длинный
тонкий трубчатый
отросток
нервной
клетки –
выходит из спинного
мозга,
вынюхивая,
словно
собачий нос,
кожу на
животе.
Другой аксон
вырастает,
нацеливаясь
на кожу на спине.
При
ненарушенном
развитии все
происходит
правильно:
щекочешь
спинку –
лягушка чувствует
спинку,
щекочешь
брюшко –
брюшко. Но у
лягушки Сперри
некоторые
нервные
клетки,
идущие на
“запах” кожи с
живота,
оказались на
спине, а
другие,
которые
направлялись
к коже спины, –
на брюшке.
Оказывается,
каждая нервная
клетка в
спинном
мозге имеет
особую метку,
например,
нервная
клетка для
брюшка или
нервная
клетка для
спинки, даже
если клетка
пока не
пустила свой
аксон к месту
назначения. И
клетка
находит свой
участок кожи,
где бы он ни
находился.
Если бы по спине
экспериментальной
лягушки
ползла муха,
то лягушка
почувствовала
бы, как муха внезапно
перескочила
на брюшко,
поползала там,
а потом снова
перескочила
на спинку.
Эксперименты
позволили Сперри
сформулировать
свою “хемоаффинную”
гипотезу,
согласно
которой
нервные
волокна
протягиваются
в нужных
направлениях
не благодаря
наличию
общего плана,
а в силу того,
что каждый
аксон ищет
свой пункт
конечного
назначения,
следуя индивидуальной
химической аффинности,
или
химическому
сродству.
Здесь снова
срабатывают
местные
условия и
правила.
Клетки в
изобилии
снабжены
метками,
химическими
значками,
которые
помогают им
найти партнеров.
В каждой
клетке, на
поверхности,
где клетки
контактируют
друг с
другом, есть
много скрепляющих
молекул.
"Клеточное
склеивание"
играет
важную роль в
эмбриональном
развитии
всех частей
тела. Однако
клеточные
клеи отличаются
от привычных
нам. Для нас
клей – это клей.
Им склеивают,
и все.
Молекулы
клеточного
клея гораздо
более
изощренные.
Более причудливые,
если можно
так сказать.
Если наш клей
склеивает
почти любые
поверхности,
то молекулы
клеточного
клея
приклеятся
только к
строго
определенным
молекулам.
Один из классов
таких
связывающих
молекул у
позвоночных
называется кадгеринами.
Кадгерины
включают
больше
восьмидесяти
известных сегодня
веществ. За
небольшим
исключением,
из примерно
восьмидесяти
кадгеринов
каждый
свяжется
только с такой
же молекулой.
Впрочем, оставим
клей, есть
аналогия
лучше:
детская игра,
в которой
каждому
игроку
дается метка
с названием
какого либо
животного, и
меток этих по
паре на
каждого
животного.
Игра
заключается
в том, что
дети носятся
по комнате и
издают звуки,
характерные
для своего животного,
а задача
игрока –
услышать в
этой какофонии
напарника и
извлечь его
из толпы. Кадгерины
действуют
сходным
образом. Но и
здесь не предполагается
наличие
плана:
действуют микроусловия
и локальные
правила.
Ферменты
Теперь,
увидев, как
зародышевые
листки
"играют в
оригами",
складываясь
в эмбрион,
давайте
заглянем
внутрь
клетки. Там
мы
столкнемся с
тем же
принципом самосборки
– самоизгибания
и самосвертывания,
но в
существенно
меньшем
масштабе – в
масштабе
белковой
молекулы.
Жизнь,
возникнув,
развивается
в рамках
процессов,
имеющих
отношение к
дарвиновскому
естественному
отбору генов.
Она во многом
основана на
белках или
других
молекулах,
которые,
подобно
белкам, умеют
складываться
во
всевозможные
многообразные
формы.
Белковые
молекулы
являются
виртуозами “автооригами”,
но размерный
класс их мира
гораздо
меньше, чем в
мире
эмбриональных
тканей, где
мы уже
побывали. На
показательных
выступлениях
по
использованию
локальных
условий белковые
молекулы
стали бы
безоговорочными
победителями.
Белки
представляют
собой
цепочки
мелких
молекул,
которые называются
аминокислотами.
Эти цепочки,
как и слои
зародышевых
клеток, имеют
свойство
складываться
строго
определенным
образом, но в
мире
меньшего
масштаба.
Природные
белки
состоят из
двадцати
типов
аминокислот.
Кстати, это
свойство на
других
планетах
может быть и
другим, потому
что во
Вселенной
больше
аминокислот,
но все земные
белки
выстроены
именно из
двадцати. Как
у них
получается “автооригами”?
Аминокислотные
цепочки
белков,
просто следуя
законам
химии и
термодинамики,
спонтанно,
сами собой
сворачиваются
в
определенные
трехмерные
конфигурации.
Я написал
было –
“завязываются”,
но в отличие
от миксин
(да, позволю
себе такое
сравнение)
белки не завязываются
в узлы. Когда
мы обсуждали
вирусы, мы
встречались с
трехмерными
структурами
белков,
получившими
название
третичных.
Они
получаются
путем
сложения и
сворачивания
аминокислотных
цепочек.
Каждая последовательность
аминокислот
диктует и строго
определенную
картину
свертывания.
Так что
последовательность
аминокислот
в цепочке,
сама по себе
определяемая
последовательностью
“букв” в коде
ДНК,
навязывает и
третичную
структуру
белку.
[Эта
фраза
требует
пояснения.
Аминокислотная
последовательность
определяется
генами и
только
генами. Но
трехмерная
форма
обусловлена
не только
последовательностью
аминокислот,
поэтому
здесь
зависимость
не
абсолютная,
что в
реальном
мире имеет
большое
значение. Некоторые
цепочки
аминокислот
способны формировать
альтернативные
трехмерные
структуры.
Белки,
называемые прионами,
имеют два
варианта стабильных
трехмерных
форм. Две
вариации строго
дискретны,
что означает,
что между этими
двумя нет
стабильных
промежуточных
переходов,
подобно
выключателю
света, который
может быть
только в двух
положениях – “включено”
и “выключено”.
Подобные
“мигающие белки”
могут быть
чрезвычайно
вредными, а
могут быть
полезными. В
случае с прионами
они ужасны.
Коровье
бешенство
вызывает одна
из
альтернативных
форм белка. В
мозге, в клеточных
мембранах
включен
некий необходимый
полезный
белок,
имеющий
альтернативную
форму. Эта
альтернативная
форма обычно
не встречается,
но стоит
появиться
хоть одной
молекуле, она
вызывает
цепную
реакцию:
соседние
молекулы
начинают
сворачиваться
по ее правилам
и сами
превращаются
в свою альтернативу.
Подобно
падающим
костям
домино или
распространяющимся
слухам,
альтернативный
прион
распространяется
по всему
мозгу с
ужасными
последствиями
для коровы
или для человека
с болезнью Крейтцфельдта
Якоба
либо для
овцы, если та
заболела
почесухой. Но
иногда
альтернативные
вариации
третичных
структур белков
могут быть
полезными.
Держа в уме
сравнение с
выключателем
света, мы
вспомним замечательный
пример.
Родопсин –
светочувствительный
белок в
глазах,
который
включает ретиналь
(это не белок
по своей
химической
природе), перескакивающий
от одной
стабильной
формы к
другой, когда
на него
попадает фотон.
Затем он
вновь быстро
возвращается
в первоначальную
стабильную
форму, отмеривая
таким
образом, как
счетчик,
световые
кванты. Но,
так или
иначе, мозг
регистрирует
эти переходы:
“Внимание, в
этой точке
зарегистрирован
свет”. В книге
Жака Моно
“Случайность
и
необходимость”
такие случаи
наглядно разобраны. (прим.
автора)]
А форма
третичной
структуры, в
свою очередь,
имеет
химические
следствия
исключительной
значимости.
Движения
“автооригами”
(теперь это
сворачивающиеся
и
складывающиеся
белковые
цепочки)
определяют
химические
законы. Те же
законы
определяют
углы между связывающимися
атомами.
Представьте
себе
ожерелье из
неправильной
формы
магнитов. Оно
не будет
обрамлять
прекрасную
шею прекрасным
овалом.
Вместо этого
магниты
перепутаются,
сцепляя углы
с выемками и
соединяя
одну плоскую
грань к другой.
В отличие от
белковой
цепочки,
форма магнитного
“украшения”
непредсказуема,
потому что
каждый
магнит
притягивается
другими и
может
сцепиться с
любым из них.
Но для цепочки
из белков
дело обстоит
иначе:
спонтанно
свернутая в
глобулу
аминокислотная
последовательность
ведет себя
совсем не так,
как ожерелье
из магнитов.
Нам
неизвестны
все нюансы
химических
закономерностей,
которые
склеивают
вместе те или
иные части
аминокислотной
цепи, но эти
закономерности
познаваемы в
принципе, они
в принципе
выводимы из этой
самой
последовательности.
Хорошо
известно (и
не раз
продемонстрировано),
что
определенная
последовательность
аминокислот
сворачивается
всегда одинаково
или
несколькими
альтернативными
способами.
Для эволюции
важно то, что
последовательность
аминокислот
определяется
согласно
правилам генетического
кода
последовательностью
букв
(триплетов
нуклеотидов)
того или иного
гена. И хотя
химикам не
так-то просто
предсказать,
как
отразится
мутация в
гене на форме
белка, следует
понимать, что
это
принципиально
возможно.
Любая
конкретная
мутация
изменит форму
фермента
совершенно
определенным
образом (или
создаст
набор
нескольких
альтернативных
форм). А
естественному
отбору только
это и нужно.
Ему нет нужды
знать, каким
образом эта форма
получена. И
если она
отразится на
выживании,
ген с данной мутацией
либо
выиграет,
либо
проиграет в
конкуренции
за
доминирование
в генофонде.
И какое дело,
понимаем мы
или нет, как
именно
сворачивается
данный белок?
Мы знаем,
что формы
белков
неисчислимо
разнообразны,
и знаем, что
они
предопределены
генетическими
последовательностями.
Но почему это
так важно?
Отчасти
потому, что
белки
являются
непосредственным
строительным
материалом
тела. Так,
волокнистые
белки,
например
коллаген,
соединяются в
прочные
жгуты,
называемые
связками и
сухожилиями.
Большинство
белков все же
не волокнистые,
они имеют
характерную
глобулярную
форму.
Глобулы
снабжены
мелкими
выступами
(или
зубчиками), и
их
взаиморасположение
обусловливает
ферментную
или
каталитическую
роль белков.
Катализатором
называют
химическое
вещество,
которое
ускоряет
химическую
реакцию
иногда в
миллион или
миллиард раз;
при этом сам
катализатор
остается
неизменным:
реакция
успешно
прошла, и
катализатор
снова
свободен и
снова готов к
работе.
Особый класс
катализаторов
составляют
белковые катализаторы,
или ферменты
(энзимы). Они
фантастически
капризны –
предельно
разборчивы в
выборе
химической
реакции. Или
наоборот:
химические
реакции в
клетке
разборчивы в
выборе
фермента.
Ведь многие
реакции в
клетке
настолько
медленные,
что без
помощи ферментов
от них бы не
было
никакого
толку. Но с ферментами
они протекают
очень быстро,
так что
продукт можно
выдавать на
гора.
Мне
нравится
представлять
это так. Вот
химическая
лаборатория
с сотнями
бутылочек и
колб, и в
каждой
простое или
сложное
вещество, раствор
или порошок.
Химик,
желающий
провести опыт,
снимает с
полки пару
тройку
бутылочек,
берет оттуда
щепоть
одного, каплю
другого,
смешивает в
отдельной
пробирке
(может и
подогреть) – и
реакция
начинается. А
сколько еще
могло бы
произойти
химических
реакций в
этой
лаборатории,
если бы все
имеющиеся
там вещества
не были
изолированы?
Живая
клетка
сродни
гигантской
химической
лаборатории
с огромными
запасами
химикатов. Но
они не
разделены
стеклянными
стенками. Все
перемешано,
будто
какой-то
громила
ворвался в
лабораторию
и расколотил
все колбы.
Ужасно,
правда? Было
бы ужасно,
если бы все
вещества
вступали в
реакцию друг
с другом в
любых
комбинациях.
Однако этого
не
происходит.
Или
происходит,
но с
ничтожной
скоростью. Если
только там
нет фермента.
Тогда все
полки с
бутылочками
становятся
ненужными,
ведь
вещества и
так не
собираются
вступать в реакцию,
пока не
появится
правильный
фермент.
Иными
словами, если
нужно, чтобы
вещества
А и Б
вступили в
химическую
реакцию,
можно хранить
их в
отдельных
закупоренных
бутылочках, а
можно слить
сотню разных
веществ в котел
и потом
добавить
фермент,
катализирующий
реакцию А и Б.
Следует
понимать, что
аналогия с
разгромленной
лабораторией,
конечно же,
не идеальна.
Ведь в клетке
имеется инфраструктура
из мембран,
которые
разграничивают
пространство,
где
протекают реакции.
Так что в
некоторой
степени
мембраны
исполняют
роль колб и
флаконов.
Смысл
этого
раздела
главы в том и
заключается,
что “правильность”
фермента
достигается
за счет правильной
формы, а она, в
свою очередь,
получается
за счет
определенной
генетической
последовательности
(последовательности
нуклеотидов
в гене), а
естественный
отбор работает
именно с
генами,
поддерживая
или отбрасывая
их. В “супе”,
который
наполняет
клетку, плещутся,
роятся и
вертятся
тысячи
молекул. Молекула А, может, и
желала бы
прореагировать
с молекулой
Б, но для
этого им
нужно
встретиться
и занять
правильную
позицию по
отношению друг
к другу. Это
очень
маловероятно,
если только
не вмешается
правильный фермент.
И у него,
обладающего
своеобразной
формой, есть
необходимые
выступы и
углубления
нужной
конфигурации.
У
каждого
фермента
имеется так
называемый
активный
центр (или
активный
сайт). Обычно
это зубец или
карман, форма
которого
придает
специфические
свойства
ферменту.
Слово “зубец”
здесь, наверное,
не совсем
уместно.
Пожалуй,
лучше сравнить
его с
электрической
розеткой.
Активные
центры на
поверхности
белковых
молекул подобны
розеткам, в
которые
можно
вставить только
вилки
определенного
типа.
Представим,
как с помощью
фермента
образуется
химическое
соединение
АБ, – очевидно,
путем
присоединения
молекулы
А к молекуле
Б. В одну
часть
активного
центра
фермента
идеально
вставляется
молекула А,
будто деталь
в пазл. Вторая
часть
активного
центра так же
идеально
подходит для
молекулы
Б, куда
она
прицепляется,
становясь в
позицию,
которая
требуется
для
соединения А
и Б. Выступы и
выемки
крепко
держат А и Б,
угол для
реакции
совершенно
точен –
молекулам А и
Б ничего не
остается,
кроме как
вступить в
реакцию.
Образуется
соединение
АБ,
высвобожденное
из объятий
фермента, а
сам фермент
готов снова
примерить к
своим
выемкам и
выступам
новые молекулы А и Б. В
клетке могут
быть целые
армии одинаковых
ферментных
молекул,
работающих
подобно
роботам на
конвейерной
ленте, выпускающей
продукт
клеточной
индустрии АБ.
А если в
клетке на
конвейере
заработает другой
фермент, то
пойдет
выпуск
другой продукции
– АВ, БД или ЕЖ.
Конечные
продукты будут
различными,
хотя сырье
всегда одно и
то же. А есть и
другие
ферменты –
они
занимаются не
соединением
молекул друг
с другом, а
расщеплением
молекул на
части.
Некоторые из
таких
ферментов
вовлечены в
процесс
пищеварения,
также они
являются
своего рода
биологическими
моющими
средствами.
Но так как в
этой главе
обсуждается
построение
эмбрионов, то
нас все
же
интересуют
строительные
ферменты, посредники
химического
синтеза.
Здесь у
читателей
могут
возникнуть
вопросы. Все
это
прекрасно:
зубцы и
выступы, вилки
и розетки,
активные
центры, у
которых сродство
только к
одним типам
молекул и
которые в
миллион раз
ускоряют
единственную
реакцию – да,
все это
хорошо.
Слишком
хорошо, чтобы
быть правдой.
Как
получаются
молекулы фермента
нужной формы?
Какова
вероятность
того, что
сразу
объявится
фермент, у
которого
есть и “вилки”,
и “розетки”
для А и Б,
причем эти
высокоспецифичные
неровности
сразу
поставят А и
Б напротив
друг друга,
заставив
любоваться
друг на
дружку под нужным
углом, а
потом еще и
соединиться?
Вероятность
ничтожна,
если
представлять
себе целиком
сразу весь пазл – или,
если вам
нравится
аналогия,
сразу все сети
“штепсельного
заговора”. Но
на самом деле
следует
представить
вот что:
постепенные
улучшения.
Как в других
случаях,
когда перед
нами встает
загадка
образования
сложных
структур,
нельзя
думать, что
их
совершенство
в настоящий
момент существовало
всегда.
Молекулы
ферментов
ускоряют
нужные реакции
в миллиард
раз. Это
выходит у них
благодаря
совершенной
форме,
приспособленной
для
определенной
реакции. Но
так ли нужна
такая
скорость?
Возможно, и
ускорения в
миллион раз
хватило бы.
Или в тысячу
раз. Даже
стократно. Да
что тут
говорить – и в
десять раз, и
даже в два
раза быстрее,
чем исходная
реакция. Даже
за это
улучшение естественный
отбор сразу
ухватился бы.
Работа
фермента
улучшалась
постепенно, его
форма
совершенствовалась,
меняясь от самой
простой,
почти без
зубцов, до
филигранно
подогнанных
друг к другу
деталей.
Постепенность
означает, что
на каждом
шаге, даже шажке,
происходило
какое-то
заметное
улучшение. А
заметность
его
предполагает,
что естественный
отбор должен
обратить на
него внимание,
даже если для
нас это
улучшение неразличимо.
Вот так
все и
работает.
Лучше не
придумаешь!
Клетка
является
живой
химической
фабрикой,
способной
выпускать
колоссальное
число
соединений в
любом
количестве,
стоит только
добавить
необходимый
фермент. И как
этот выбор
делается?
Путем
включения нужного
гена. Если
клетка
представляется
котлом с
множеством
химикатов, то
в клеточном
ядре,
напротив,
работает
только малое
число генов.
Если ген
включен
(например, мы
говорим о
клетке
печени), то
последовательность
нуклеотидов,
то есть
генетический
код, определит
последовательность
аминокислот,
она
определит
форму, в
которую
свернется белок,
а в этой
форме
появятся
нужные зубцы
и впадины,
которые
неизбежно
заставят
молекулы в
“котле”
соединиться
требуемым
образом. В каждой
клетке, за
исключением
красных кровяных
телец,
имеется ядро,
содержащее
набор генов
для всех
ферментов. Но
также в каждой
клетке
“активированы”
лишь
немногие
гены. Так, в
клетках
щитовидной
железы
вырабатываются
ферменты для
производства
тироидного
гормона, и
соответственно,
работают
только гены
этих
ферментов.
Точно так и с
любыми другими
клетками.
Конфигурация
клетки, ее поведение,
ее участие в
спектакле –
все определяется
идущими в
клетке
химическими
реакциями.
Поэтому весь
ход
эмбрионального
развития
управляется
генами.
Именно гены диктуют
последовательность
аминокислот, определяющую
третичную
структуру
белков,
именно она
формирует
“розетки” и “штепсели”
в активном
центре. Он, в
свою очередь,
контролирует
клеточную
химию, а это
вызывает
спонтанно
организованное,
подобное
поведению
скворцов в
стае,
поведение
клеток в
процессах эмбрионального
развития.
Таким
образом, разница
в генах может
вызвать
изменения хода
эмбрионального
развития и,
следовательно,
изменения
облика и
поведения
взрослых
особей. А
впоследствии
выживание и
репродуктивный
успех этих
особей
повлияют на
выживание
генов,
которые
некогда
обеспечили
их выживание
и
репродуктивный
успех. И в
этом заключается
естественный
отбор.
Эмбриональное
развитие
кажется
сложным (так
оно и есть), но
все видится
проще, если
разложить
его на более
простые
составляющие,
которые, по
сути, являются
самоорганизующимися
процессами,
управляемыми
во всех
случаях
местными
взаимодействиями.
Другое дело –
понять, как
клетки, в
которых
содержится
набор генов,
решают, какой
из генов
следует
задействовать
в определенный
момент. Попробуем
ответить и на
этот вопрос.
Утеха
для червей
Будет ли
тот или иной
ген
активирован,
зависит от
химического
состава
окружающей
среды, а он регулируется
работой
каскадов
других генов,
называемых
генами
переключателями
либо
регуляторными
генами.
Клетки
щитовидной
железы,
например,
сильно
отличаются
от мышечных,
как и от всех
других,
понятное
дело, хотя
гены у всех
одинаковые.
Хорошо, скажете
вы: клетки
мышц и
щитовидной
железы
различаются
у более или
менее
развившихся
эмбрионов. Но
эмбрион
начинается с
одной
единственной
клетки.
Клетки
щитовидной и
поджелудочной
желез, клетки
печени и
костей,
клетки кожи и
мышц – все это
потомки
оплодотворенной
яйцеклетки,
их судьбы
складываются
в ветвящееся
генеалогическое
древо. Это
древо, берущее
начало в
зачатии, не
имеет ничего
общего с
эволюционным
древом,
уходящим в
прошлое на
миллионы лет,
которым мы
любовались в
других
главах.
Позвольте
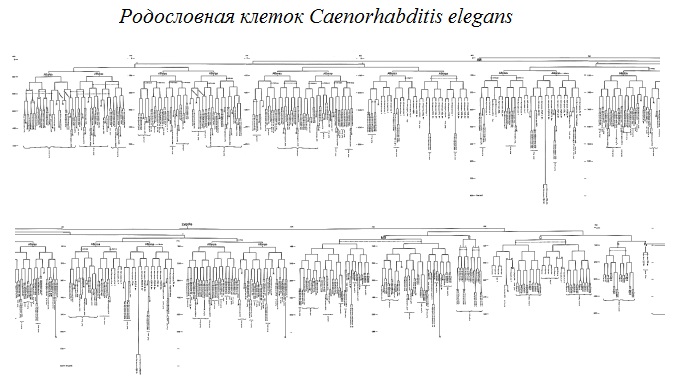
показать вам,
например,
полную
родословную
558 клеток
только что
вышедшей из
яйца личинки
червя
нематоды Caenorhabditis elegans.
Не знаю,
почему этим
червям дали
видовое название
elegans –
изящный, но
теперь можно
сказать, что
они его
заслуживают.
И, хотя я знаю,
что не все
читатели
одобряют
отступления
от темы,
исследования
C. elegans –
это такое
торжество
научной
мысли, что сейчас
меня ничто не
остановит!
В 1960-х
блистательный
южноафриканский
ученый
Сидней Бреннер
выбрал C. elegans в
качестве
идеального
подопытного
животного. Он
как раз
закончил
свою работу с
Фрэнсисом
Криком и
другими
коллегами из
Кембриджа, посвященную
расшифровке
генома, и
обдумывал, за
что бы
взяться. С
его легкой
руки – и благодаря
его мощному
творческому
заделу! – в исследованиях
генетики и
анатомии
нервной
системы C. elegans
появилось
целое
сообщество
специалистов
по C. elegans,
которое
теперь
насчитывает
тысячи людей.
Можно
сказать, что
мы знаем о C. elegans почти все.
Нам
полностью
известен его
геном. Мы
точно знаем,
где
находится
каждая из его
558 клеток (у
личинок; у
взрослой
особи
гермафродита
их 959, не считая
репродуктивных),
и для каждой
составлена
полная родословная.
Нам известно
большое
количество
мутантных
генов,
приводящих к
появлению атипичных
червей, и мы
точно знаем,
где в теле
проявляется
мутация. Мы
знаем и полную
историю
образования
этих
отклонений.
Это
маленькое
создание
известно нам
снаружи и
изнутри,
вдоль и
поперек. В 2002
году Бреннер
был, пусть
запоздало,
удостоен
Нобелевской
премии, и
один из
видов,
близких к C. elegans,
был назван в
его честь – Caenorhabditis brenneri.
Его
постоянная
колонка “Дядя
Сид” в журнале
“Каррент байолоджи”
являет
образец
эрудированности
и научной
мудрости,
остроумной и
прекрасной,
как вся
область
исследований
C. elegans,
им
инспирированная.
Однако мне
хотелось бы,
чтобы
микробиологи
общались с
зоологами
(как это
делал сам Бреннер)
и перестали
бы
относиться к Caenorhabditis
как к
единственной
нематоде или
даже как к единственному
червю, будто
других не
существует.
Разумеется,
в диаграмме
вы не сможете
прочесть, где
какие типы
клеток
находятся
(чтобы
написать их
разборчиво, понадобилось
бы семь
страниц), но
там указано:
“глотка”,
“мышцы
кишечника”,
“мышцы тела”, “сфинктер”,
“кольцевой
ганглий”,
“поясничный ганглий”.
Все эти
клетки вне
зависимости
от типа
приходятся
друг другу
буквально
двоюродными
сестрами с
учетом их
общего происхождения.
Я
рассматриваю
мышечную клетку
MSpappppa.
Она является
сестрой
другой
мышечной
клетки,
двоюродной
сестрой двух
мышечных
клеток,
двоюродной
племянницей
еще двух
мышечных
клеток,
троюродной –
шести клеткам
глотки,
четвероюродной
сестрой еще семнадцати
клеткам
глотки, и так
далее. Не правда
ли
поразительно,
что мы можем
уверенно
использовать
слова вроде
“племянница”
по отношению
к клеткам,
каждая из
которых поименована
и однозначно
определена?
Количество
“поколений”
клеток,
отделяющих
ткани от первоначальной
яйцеклетки,
невелико. В
конце концов,
в теле C. elegans всего
558 клеток, а
теоретически
можно получить
1024 (два в
десятой
степени)
клеток всего
за десять
делений.
Количество
поколений
клеток у
человека
гораздо
больше. Тем
не менее,
теоретически
можно
составить
аналогичную
родословную
каждой из
триллионов
клеток
(сравните с 558
клетками
женской
личинки C. elegans),
проследив
линию
предков до
одной
оплодотворенной
яйцеклетки.
Впрочем, для
млекопитающих
невозможно
определить и
поименовать
конкретные
клетки, и так,
чтобы это
соответствовало
их
расположению.
У нас речь
идет о некой
усредненной
популяции
клеток,
немного различающихся
у разных
людей.
Надеюсь,
что моя ода
во славу Caenorhabditis
не увела нас слишком
далеко от
разговора о
том, как различные
клетки
отдаляются
друг от
друга, меняя
форму и
другие
признаки по
мере ветвления
древа клеток
в ходе
эмбрионального
развития. В
том месте,
где двум
линиям
клеток суждено
разойтись в
разные
стороны,
должно произойти
нечто, что
будет
отличать
клетку, чья
судьба –
стать
клеткой
гортани, от
ее сестры
близнеца,
которая
превратится
в кольцевой
ганглий. А
иначе как они
определят,
какие гены
какой клетке
задействовать?
Дело в том,
что когда
ближайший
общий предок
двух клонов
делился
пополам, две
половины его клетки
уже не были
одинаковыми.
Поэтому две
дочерние
клетки, хотя
и получили
одинаковый
набор генов,
различались
по химическому
составу.
Вследствие
этого у них
были активированы
разные гены,
что и
определило
дальнейшую
судьбу их
потомков.
Этот принцип
действует на
протяжении
всего
развития
эмбриона, с
самого
начала. Ключ
к клеточной
дифференциации
у всех
животных –
асимметричное
деление
клеток.
У C. еlegans
первая
клетка
обозначается
Z
(“зигота”), и ее
передний конец
отличается
от заднего.
Это различие
реализуется
в итоге как передне задняя
ось тела.
Когда
делится
зигота, то
передняя
дочерняя
клетка,
называемая AB,
получает
больше “передне
задней”
субстанции,
чем задняя
дочерняя клетка
Р1.
Различие
между
линиями этих
клеток в
дальнейшем
усиливается.
АВ дает начало
большей
части клеток
тела, включая
почти всю
нервную
систему, и
эту часть я
больше не
буду
обсуждать. Р1 рождает
двух дочек,
опять же не
сходных друг
с дружкой: EMS
(она дает
начало
остальным
клеткам
брюшной
стороны) и Р2
(отвечает за
клетки
спинной стороны).
Обе являются
внучками
зиготы Z (не
забывайте,
что мы
обсуждаем
потомство клеток,
а не
отдельных
нематод). EMS
делится
пополам, и
двоих ее
отпрысков
называют E и MS, а
отпрысков Р2 –
С и Р3. Эти
четверо являются
правнучками Z
(правнучки АВ
также
существуют,
но я о них не
упоминаю, за
исключением
потомков ABal
и ABpl,
определяющих
левую
сторону, и их
двоюродных
сестер ABar и ABpr,
определяющих
правую). У Р3
появляется
две дочки D и P4 –
это
праправнучки
Z. MS
и С также
имеют
потомков,
каждого со
своим именем.
Потомство P4
превращается
в так
называемую
зародышевую
линию. Клетки
зародышевой
линии не участвуют
в
формировании
тела червя,
они развиваются
в
репродуктивные
клетки.
Понятно, что
запоминать
все эти названия
не нужно.
Важно только
то, что хотя они
все
генетически
идентичны,
химическое
содержание у
них
различается. Причина
этих различий
кроется в
индивидуальной
истории
клетки.
Сэр Джон
Салстон
с коллегами
проследил
путь каждой
из клеток
тела червя
вплоть до
одной из
шести клеток
основательниц.
Мы можем
назвать их
клетками
родоначальницами:
AB, MS, E, D, C и P4. [Салстон,
оставшийся в
Кембридже
после того,
как Бреннер
уехал в
Америку, стал
еще одним
нобелевским
лауреатом,
получившим
премию за
работы по C. elegans.
Впоследствии
он возглавил
британскую
ветвь
проекта
“Геном
человека”,
который в
Америке
возглавлял
сначала
Джеймс
Уотсон, а после
Фрэнсис Коллинз. (пример
автора)]
Когда
ученые
давали
клеткам
названия, они
стремились
сделать так, чтобы
история каждой
была сразу
ясна.
Название
клетки начинается
с названия
одной из
шести клеток
родоначальниц.
Дальнейшее
название
клетки
состоит из
букв,
указывающих
направление
деления:
перед (anterior),
зад (posterior),
спина (dorsal),
брюхо (ventral),
лево (left) или
право (right).
Например, Ca
и Cp – две
“дочери”
клетки
родоначальницы
C (передняя и
задняя
соответственно).
Обратите внимание:
у каждой
клетки может
быть не более
двух
“дочерей”
(одна может
погибнуть).
Вот мышечная
клетка Cappppv.
У нее
говорящее
имя. У клетки C
была
передняя
дочерняя
клетка, от которой
отделилась
задняя “дочь”,
у которой также
была задняя
“дочь”, и у двух
следующих также.
Последняя
породила
брюшную
дочернюю
клетку, на
которую я
смотрю.
Каждая клетка
организма
описана
строчкой,
заглавные
буквы
которой
являются
именем одной
из клеток
основательниц.
Еще один
пример –
клетка ABprpapppap,
нервная
клетка,
расположенная
в брюшном нерве,
тянущемся по
всей длине
червя. Наверное,
нет
необходимости
перечислять
все ее деления.
Замечательно
то, что у
каждой
клетки есть
имя,
описывающее
историю ее
развития.
Каждое из
десяти
делений, которые
привели к
появлению
клетки ABprpapppap
(понятно, не
только ее, но
и остальных),
было асимметричным,
что
предопределило
активацию
разных генов
для двух
дочерних
клеток. Таким
же образом
происходит
дифференциация
тканей во
всех живых
существах.
Разумеется, у
большинства
животных
больше 558 клеток,
и их
эмбриональное
развитие, как
правило,
задано менее
жестко. Как по
дружески
напомнил мне
сэр Джон Салстон
(и о чем я уже
коротко
упоминал), у
млекопитающих
(особенно у
них)
клеточные
родословные
различны для
каждой особи,
а у Caenorhabditis они
практически
идентичны,
исключая
мутантных
особей. Тем
не менее,
принцип
остается тем
же. У любого
животного
клетки
различных
частей тела,
хотя и несут
одинаковый
генетический
код,
отличаются
друг от друга
именно из-за
асимметричного
деления в
процессе
короткой
эмбриональной
истории.
Подведем
итоги. Не
существует
никакого плана
развития,
никакой
схемы,
никакого
чертежа, да и
самого проектировщика
также нет.
Развитие
эмбриона, а в
итоге и всего
организма,
определяется
набором
локальных
механизмов,
применяемых
к отдельным
клеткам,
взаимодействующих
в местном
масштабе
здесь и
сейчас.
Внутриклеточные
процессы
также
регулируются
локальными правилами,
применяемыми
к отдельным
молекулам,
особенно
белкам.
Повторяю: все
эти механизмы
и правила
локальны –
только “здесь
и сейчас”!
Теперь,
чтобы эта
глава не
выглядела
отступлением
от главной
темы книги –
эволюции, вернемся
к Холдейну
и
скептически настроенной
даме. Если
гены
определяют
пути
развития
эмбриона, а
не форму
взрослого организма,
если ни
естественный
отбор, ни Бог,
в отличие от
процессов
эмбриогенеза,
не "изготавливают
крылышки", то как
действует
естественный
отбор у животных,
когда формирует
их тела и
поведение?
Как
естественный
отбор
действует на
эмбрионы,
сортируя и вытачивая
детали,
предоставляя
взрослым животным
именно то,
что им
необходимо
для выживания:
крылья или
плавники,
щитки или
крепкую
броню, жало
или щупальца?
Естественный
отбор – это
выборочное
выживание
успешных генов
по отношению
к другим,
менее
успешным. Но
естественный
отбор не
выбирает
гены напрямую.
Вместо этого
он отбирает
их носителей
– отдельных
особей. Этот
выбор
происходит
сам собой.
Ненамеренно
отбираются
те, кто сумел
выжить и
оставить
потомство,
передав ему
те самые
гены.
Выживание
гена зависит от
выживания
организма, на
образование
которого
повлиял этот
ген: ген,
будто
всадник, едет
вместе с
телом, они
вместе живут
и вместе
умирают.
Любой ген
может
существовать
одновременно
в виде
множества
копий, путешествующих
в большом
числе тел в
поколении, и
передаваться
из поколения
в поколение в
популяции. Но
количества
копий гена,
который в
среднем
увеличивает
шансы на
выживание,
будет в
популяции
постепенно
увеличиваться.
Таким
образом,
гены, которые
мы встречаем
в генофонде,
скорее всего,
будут теми
генами, которые
формируют
тела более
успешным образом.
В этой
главе мы
увидели, как
именно гены
формируют
тело. Дама,
задавшая
вопрос Холдейну,
полагала, что
естественному
отбору вряд
ли хватило бы
миллиарда
лет, чтобы
создать
генетический
рецепт
изготовления
человека. Я
же утверждаю,
что это вероятно.
Хотя,
разумеется,
ни я, ни кто бы
то ни было
еще не может
подробно
рассказать,
как это
происходило.
Это оттого,
что все правила
имеют
локальный
характер.
Любой акт естественного
отбора
отбирает
мутацию, которая
сказалась
одновременно
на большом количестве
клеток
многих
особей в виде
небольшого
изменения
формы белка.
Он, в свою очередь,
ускоряет
какую-нибудь
химическую реакцию
во всех
клетках, в
которых
активирован
данный ген.
Это,
например,
увеличивает
скорость роста
зачатка
челюсти у
эмбриона.
Впоследствии
это
отразится на
форме головы
(к примеру,
укорачивается
морда,
черты
становятся
скорее
человеческими,
чем
обезьяньими).
Естественный
отбор может
оказывать давление
каким угодно
образом.
Например,
через эстетическое
предпочтение
потенциальных
половых
партнеров.
Изменение
формы
челюсти может
оказать
небольшое
влияние на
способность
грызть орехи
или кусаться.
Комбинация
давлений естественного
отбора,
конфликтующих,
находящих
компромиссы
невообразимой
сложности,
определяет
статистический
успех данного
гена,
пытающегося
распространиться
в генофонде.
Сам же ген не
подозревает
ни о чем.
Единственное,
что он
делает,
передаваясь
из поколения
в поколение, –
это изменяет
аккуратную
впадинку на
белковой
молекуле. Все
остальное –
неизбежное
следствие
небольших
каскадных
изменений
локального
масштаба, которые
в итоге
образуют
организм.
Но еще
сложнее, чем
сложение
векторов
отбора,
действующих
в рамках
экологических,
половых,
социальных
законов,
устроена
невообразимо
сложная сеть
взаимодействий,
реализующихся
внутри и
между
развивающимися
клетками:
влияние
генов на
белки, генов
друг на
друга, влияние
белков на
экспрессию
генов, одних
белков на другие,
мембран,
химических
градиентов,
физических и
химических
потоков
внутри эмбриона,
гормонов и
других
посредников
для управления
на
расстоянии,
клеток, нацеленных
на клетки с комплементарными
маркерами.
Никто не
может
охватить
картину
целиком, но
для того,
чтобы
принять
действенность
естественного
отбора, это и
не требуется.
Естественный
отбор
обеспечивает
распространение
в генофонде
мутаций,
которые
отвечают за
важные
изменения в
эмбриональном
развитии.
Картина
складывается
из сотен
тысяч мелких
локальных
взаимодействий,
в принципе
понятных (хотя
иногда на
практике
очень сложно
перепутанных)
любому, кто
возьмет на
себя труд разобраться.
Целое
ошеломляет,
кажется таинственным,
но на самом
деле тайны
нет. Нам известны
и принципы
эмбриологии,
и механизмы эволюционной
истории,
ведущей к
доминированию
в генофонде
определенных
генов. В процессе
эволюции
усложнения
постепенно накапливались.
Каждый шаг
уводил
немного дальше.
И каждый шаг
совершался
за счет небольшого
изменения
локальных
правил. Когда
имеется достаточное
число
элементов
(клеток,
белковых
молекул,
мембран),
подчиняющихся
локальным
правилам и
влияющих
друг на
друга, результат
может быть
впечатляющим.
Собеседница
Холдейна
ошибалась. В
этом нет
ничего
принципиально
невозможного.
И, как сказал
ей Холдейн,
на это нужно
всего девять
месяцев.
Всю
книгу, а
также и
другие
книжки Докинза,
можно за
копейки купить
тут: http://www.litres.ru/richard-dokinz/
#Biology
#Embryology
#Evolution